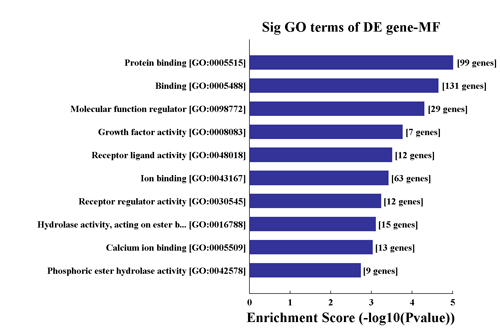
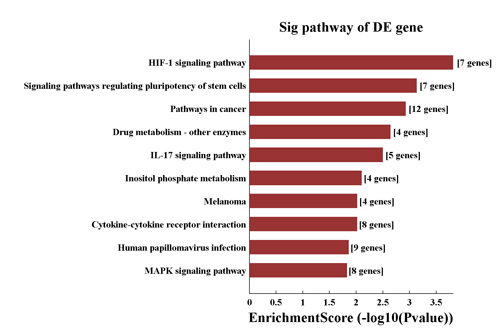
Identification and characterization of a novel adiponectin receptor agonist adipo anti-inflammation agonist and its anti-inflammatory effects in vitro and in vivo. Qiu W, et al. British Journal of Pharmacology, 2021
A surrogate of Roux-en-Y gastric bypass (the enterogastro anastomosis surgery) regulates multiple beta-cell pathways during resolution of diabetes in ob/ob mice. Amouyal C, et al. EBioMedicine, 2020
Dietary carbohydrates modulate metabolic and beta cell adaptation to high fat diet-induced obesity. Her T, et al. American Journal of Physiology-Endocrinology and Metabolism. 2020
Astrocytes influence medulloblastoma phenotypes and CD133 surface expression. Gronseth E, et al. PloS one, 2020
The Toxoplasma effector TEEGR promotes parasite persistence by modulating NF-?B signalling via EZH2. Braun L, et al. Nature microbiology, 2019
Application of hydroxyapatite nanoparticles in tumor-associated bone segmental defect. Zhang K, et al. Science Advances, 2019
Engineered triple inhibitory receptor resistance improves anti-tumor CAR-T cell performance via CD56. Zou F, et al. Nature communications, 2019
Lymphocyte-Derived Exosomal MicroRNAs Promote Pancreatic ß Cell Death and May Contribute to Type 1 Diabetes Development. Guay C,et al. Cell metabolism, 2018
Osteoblast-secreted factors mediate dormancy of metastatic prostate cancer in the bone via activation of the TGFßRIII-p38MAPK-pS249/T252RB pathway. Yu-Lee L Y, et al. Cancer Research, 2018
MicroRNA-Mediated Dynamic Bidirectional Shift between the Subclasses of Glioblastoma Stem-like Cells. Rooj A K, et al. Cell Reports, 2017
MicroRNA Signatures and Molecular Subtypes of Glioblastoma: The Role of Extracellular Transfer. Godlewski J, et al. Stem Cell Reports, 2017
Bmal1 is required for beta cell compensatory expansion, survival and metabolic adaptation to diet-induced obesity in mice. Rakshit K, et al. Diabetologia, 2016
FOXP3 promotes tumor growth and metastasis by activating Wnt/ß-catenin signaling pathway and EMT in non-small cell lung cancer. Yang S,et al. Molecular Cancer, 2017
Long noncoding RNA CRNDE stabilized by hnRNPUL2 accelerates cell proliferation and migration in colorectal carcinoma via activating Ras/MAPK signaling pathways. Jiang H, et al. Cell Death & Disease, 2017
In vitro expansion impaired the stemness of early passage mesenchymal stem cells for treatment of cartilage defects. Jiang T, et al. Cell Death & Disease, 2017
Circular RNAs as novel regulators of ß-cell functions in normal and disease conditions. Stoll L, et al. Molecular Metabolism, 2017
More Publications>>