
m6A is the most prominent modification in mRNA epitranscriptomics. Similarly, m6A modification is also abundant in amount, common in occurrence, and functionally important in the long non-coding RNA (lncRNA) epitranscriptome (Fig.1) [1, 2].
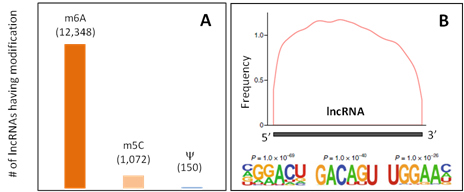
Figure 1. (A) The numbers of lncRNA transcripts having m6A, m5C or Y modifications. (B) m6A modification frequency along the lncRNA length and m6A site motifs.
The “m6A switches”
m6A modifications in lncRNAs can often form a “m6A switch”, where poly-U5 in the RNA stem-loop structure is opened or closed by m6A modification for hnRNP-C reader binding (Fig. 2)[3,4]. m6A switches are a common mechanism for readers without a YTH-domain, such as hnRNPs, where the reader does not bind directly to m6A but its adjacent poly-U5 . So far, about 39,000 m6A switches are known. In lncRNA-Malat1, m6A switches affect its RNA splicing and abundance, controlling cell cycle and lung cancer activation.
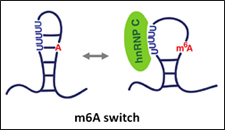
Figure 2. m6A modification weakens the base paring in the stem-loop, opening the poly-U5 to hnRNP-C biding as the reader. m6A can thus switch the hnRNP-C binding on/off [3,4].
m6A as the regulatory and functional component of lncRNAs
LncRNA m6A modification recognized by readers, such as YTH family proteins, can directly modulate the lncRNA molecular functions. For example, when lncRNA XIST is modified by METL3, WTAP, RBM15/15B writers, YTH-DC1 readers recruit chromatin silencing proteins to condense and inactivate the X-chromosome [5].
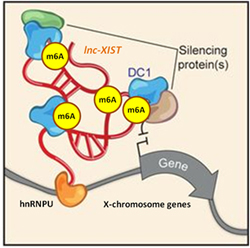
Figure 3. m6A modifications in lncRNA XIST are recognized by YTH-DC1 reader, which recruits chromatin silencing proteins to inactivate X-chromosome [5].
Many lncRNAs in the cytoplasm can act as competing endogenous RNAs (ceRNA), which competitively sponge silencing microRNAs away from the target mRNAs [6]. For linc1281, m6A modification is required for efficient microRNA let-7 binding to the m6A site (Fig. 4). With the m6A in linc1281, the let-7 target mRNA LIN28 is protected from microRNA silencing and up-regulated to promote proper cell differentiation [7].
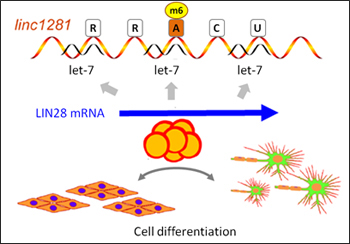
Figure 4. m6A modified lncRNA linc1218 efficiently binds let-7 microRNA, preventing let-7 from silencing LIN28 mRNA. The increased LIN28 leads to cell differentiation. Without the m6A, the cell differentiation is restricted [7].
m6A as the epitranscriptomic mark for RNA decay
As in mRNA, m6A modification can mark the lncRNAs for decay and rapid turnover. In dendritic cells, for example, m6A modification at two sites in lncRNA Dpf-3 can leads to its decay and down-regulation (Fig. 5) [8]. Without the lnc-Dpf3 inhibiting HIF-1a, the dendritic cells migrate from skin into lymph nodes for activated immune responses. With chemokine CCL19/21 activated receptor CCR7 to demethylate lnc-Dpf3, the dendritic cell immune response is terminated.
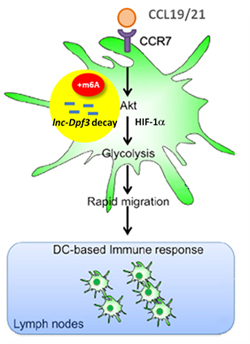
Figure 5. m6A modification of lncRNA-Dpf3 leads to its decay [8]. Without the inhibition of HIF-1a by lnc-Dpf3, the dendritic cells migrate to lymph nodes to activate immune response. With chemokine CCL19/21 stimulation, lnc-Dpf3 is demethylated and stabilized, leaving HIF-1a to terminate the dendritic cell based immune response.
Related Service
LncRNA Epitranscriptomic Array Service
References
[1] Jacob, R., et al. (2017) Int J Mol Sci [PMID: 29125541]
[2] Shafik, A., et al. (2016) Biochim Biophys Acta [PMID: 26541084]
[3] Liu N. et al. (2015) Nature [PMID: 25719671]
[4] Liu N. et al. (2013) RNA [PMID: 24141618]
[5] Patil D.P. et al. (2016) Nature [PMID: 27602518]
[6] Tay, Y., et al. (2014) Nature [PMID: 24429633]
[7] Yang D. et al. (2018) Nucleic Acids Res. [PMID: 29529255]
[8] Liu J. et al. (2019) Immunity [PMID: 30824325]