
Overview
Transfer RNA (tRNA) is an adaptor molecule that decodes mRNA and translates protein. Recent studies have demonstrated that tRNAs also serve as a major source of small non-coding RNAs having distinct and varied functions [1]. These tRNA-derived ncRNAs are not random degradation products, but rather generated through precise biogenesis processes (Fig. 1). tRNA-derived ncRNAs can be broadly classified into two main groups: tiRNAs (or tRNA halves) and tRFs (tRNA-derived fragments), with characteristic sizes, nucleotide compositions, functions and biogenesis [1-3].
tRNA halves (tiRNAs) are produced by specific cleavage in the anticodon loop of mature tRNA by angiogenin under various stress conditions to produce 29-50 nucleotide 5’-tRNA and 3’-tRNA halves.
tRFs are 16~28 nt fragments derived from tRNA or pre-tRNA, which are classified by their sites of origin: (i) tRF-5 derived from the 5’ parts of mature tRNAs by cleavage in the D loop. (ii) tRF-3 derived from the 3’ part of mature tRNAs by cleavage at the T loop. The 3’end contains a 3’-CCA terminus. (iii) tRF-1 originated from the 3’trailer from the pre-tRNA. The 3’end has poly-U residues at the end. (iv) i-tRF, which does not belong to tRF-5, tRF-3 or tRF-1, is typically derived from the internal region of the mature tRNA.
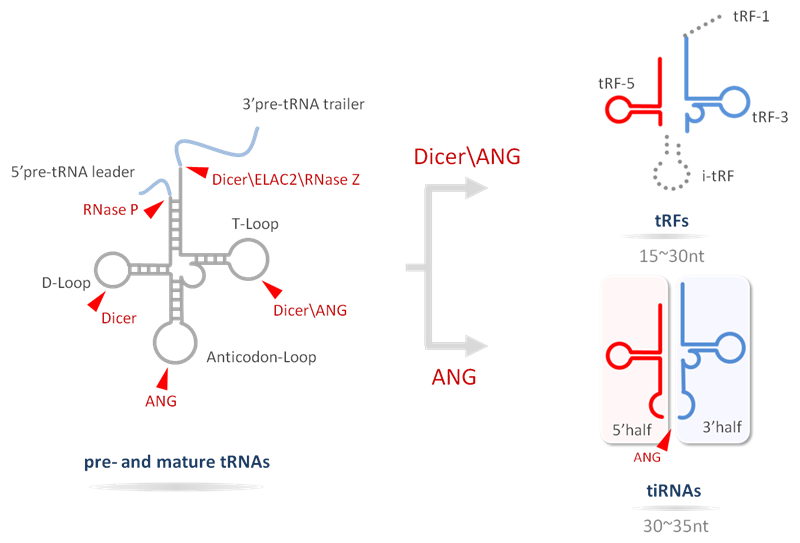
Figure. 1 Biogenesis of tRFs/tiRNAs. tRF-1 is generated from the 3’-trailer of primary tRNA. tRF-5, i-tRF and tRF-3 are produced from the 5’-, internal- and 3’-portions of the mature tRNA, respectively. When the cleavage site is within the anticodon loop, two fragments are generated as tiRNAs for the 5’- and 3’- tRNA halves.
Biological functions
tRFs and tiRNAs perform many biological functions as small noncoding RNAs (Fig.7). They are known to act as microRNAs in RNA interference (Fig. 2); directly inhibit protein synthesis by displacing eIF4G translation initiation factor eIF4G from mRNA on ribosomes [9-10]; bind protein factors such as CBX1 to regulate target mRNA stability (Fig. 3); interact with cytochrome c to modulate apoptosis [13]; assemble stress granules in response to stress conditions (Fig. 4); sensitize cells to oxidative-stress-induced p53 activation and p53-dependent cell death [19]; alter transcriptional cascades in intergenerational inheritance as paternal epigenetic factors (Fig. 6).
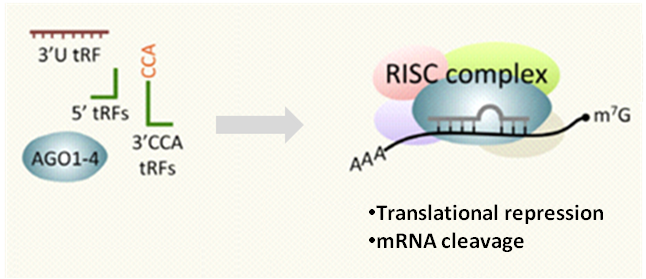
Figure 2. tRFs have many functional characteristics of a microRNA, such as Dicer-dependent biogenesis, RISC complex formation with Argonaute proteins, and RNA silencing. Some of the catalogued miRNAs directly map to tRFs [8].
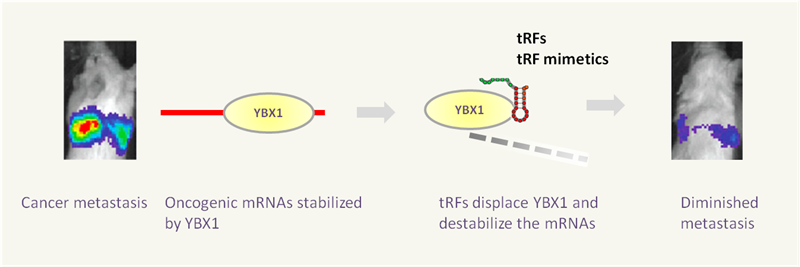
Figure 3. tRFs or the mimetics displace oncogenic RNA binding protein YBX1 and destabilize many cancer driver mRNAs. Consequently, the cancer metastasis is greatly diminished [11].
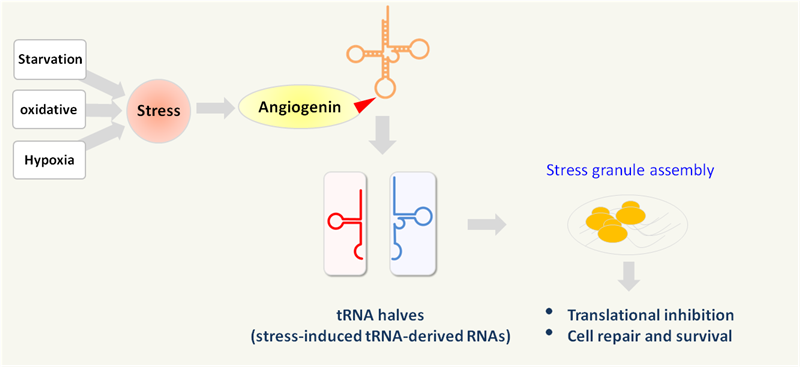
Figure 4. tRNA halves (tiRNAs) are generated by angiogenin cleavage in response to cellular stresses, which promote stress granule assembly and induce translational inhibition, cell-repair and survival [12].
Human Diseases
tRF/tiRNAs are associated with or are causal factors for disease conditions including cancers, neurodegeneration, and metabolic disorders (Fig. 5).
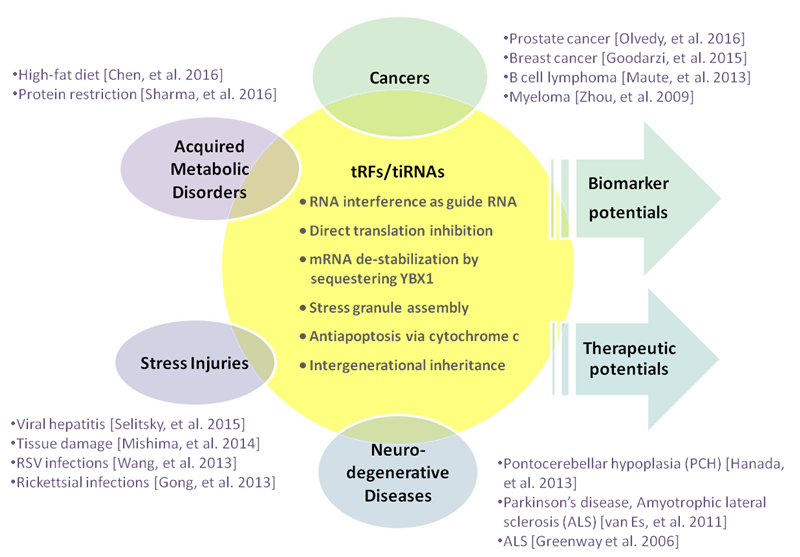
Figure 5. tRF/tiRNA molecular functions and diseases.
Cancers
Differential expression of tRFs/tiRNAs is detected in different cancer cell lines, including the prostate cancer cell lines LNCaP and C4-2. tRFs/tiRNAs levels are elevated by cellular stress conditions and particularly under oxidative stress such as hypoxia [16]. Moreover, a 3’-derived tRF identified in B-cell lymphoma cells possesses the functional characteristics of a guide RNA that suppresses proliferation and modulates response to DNA damage in a miRNA-fashion [7]. By competing YBX1 binding sites and destabilization of oncogenic transcripts, tRFs/tiRNAs act as tumor suppressors [11] (Fig. 3). tRF-1001, a 3’tRF derived from pre-tRNA-Ser, is highly expressed in different cancer cell lines, and is required for proliferation of prostate cancer cells [21]. Additionally ANG produced tiRNAs promote the assembly of stress granules that help cells to survive under adverse conditions. It is possible that ANG-induced tiRNAs directly contribute to ANG-mediated angiogenesis and cancer cell proliferation. Similarly, tiRNAs can help cancer cells to prevent apoptosis by binding to Cyt c [13]. Taken together, these findings strongly suggest a functional role of tRFs/ tiRNAs in tumorigenesis.
Acquired metabolic disorder
Increasing evidence indicates that offspring metabolic disorders can result from the father’s diet. In a paternal high-fat diet (HFD) mouse model, a subset of sperm tRFs/tiRNAs, mainly from 5’-halves and ranging in size from 30 to 34 nucleotides, exhibit changes in expression profiles and RNA modifications under high fat diet condition. Injection of sperm tRFs/tiRNAs fractions from HFD male into normal zygotes generated metabolic disorders in the F1 offspring and altered gene expression of metabolic pathways in early embryos and islets of F1 offspring, which was unrelated to DNA methylation at CpG-enriched regions. Hence, sperm tRFs/tiRNAs represent a type of paternal epigenetic factor that may mediate intergenerational inheritance of diet-induced metabolic disorder [14] (Fig.6 ).
Protein restriction in mice also affects small RNA levels in mature sperm, with decreased let-7 levels and increased levels of 5’- tRFs/tiRNAs of glycine tRNAs. tRFs/tiRNAs have been linked to the regulation of expression of transcripts driven by endogenous retroelements active in the preimplantation embryo [22].
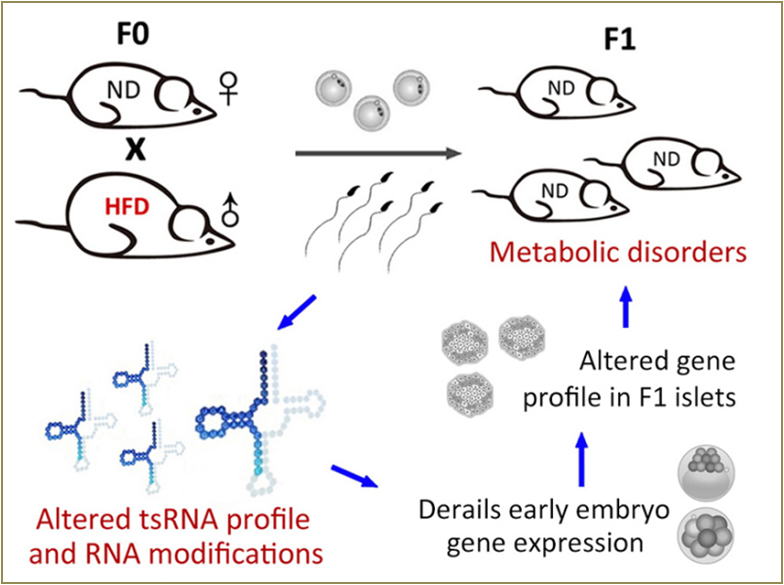
Figure 6. tRNA-derived small RNAs (tsRNAs, mostly tRNA halves) have altered expression profiles and RNA modifications in the sperms of high fat diet fed mice. The sperm tsRNAs confer the metabolic disorder phenotypes in the offspring by oocyte injection. This tsRNA paternal epigenetic factor mediates intergenerational inheritance by modulating embryonic gene transcription that cascades into the adulthood [14].
Neurological disorders
A number of neurological disorders are caused by defects in tRNA metabolism and tRNA processing enzymes (e.g. ANG). Angiogenin mutants with reduced RNase activity are implicated in the pathogenesis of Amyotrophic Lateral Sclerosis (ALS), a fatal neurodegenerative disease [23]. A subset of ALS-associated ANG mutants was also found in Parkinson’s Disease (PD) patients [24]. Further links are strengthened between ANG-induced tRNA halves, cellular stress and neurodevelopmental disorders [25]. Loss of RNA kinase CLP1 activity results in accumulation of a novel set of small RNA fragments, derived from aberrant processing of tyrosine pre-tRNA. These tRFs/tiRNAs sensitize cells to oxidative-stress-induced p53 activation and p53-dependent neuronal cell death, leading to motor neuron loss, muscle denervation and respiratory failure [19]. Another example is the mutations in the cytosine-5 RNA methyltransferase NSun2 that cause microcephaly and other neurological abnormalities. Loss of cytosine-5 RNA methylation of tRNA in the absence of NSun2 increases the ANG binding and cleavage of tRNA leading to an accumulation of 5′-halves, which reduces protein translation rates and activates stress pathways leading to reduced cell size and increased apoptosis of cortical, hippocampal and striatal neurons [25].
Pathological stress injuries
Stress imparted by hypoxia, nutrient deprivation, oxidative conditions and metabolic imbalance can damage cells and promote disease. These stresses stimulate production of tRNA halves. In animal models of tissue damage (e.g., toxic injury, irradiation and ischemic reperfusion), the production of tRNA halves is correlated with the degree of tissue damage due t, for example, a change in tRNA conformation that promotes ANG-mediated production of tRNA halves [1]. Increased expression of 5’-halves has been associated with viral and rickettsial infections, and may serve to prevent apoptosis and promote cell survival. tRFs/tiRNAs, specifically 30-35 nucleotide long 5′-halves, are abundant in non-malignant liver and significantly increased with chronic viral hepatitis [27].
Biomarker potentials
The composition and abundance of tRFs/tiRNAs are highly dependent on the cell type and disease condition [15]. In particular, tRNA and tRF/tiRNA populations are highly enriched in biofluids, sometimes more so than microRNAs [16-18] (Fig. 7A). Although miRNAs have been current main focus as candidate biofluid-based biomarkers, the high stability and abundance of tRFs/tiRNAs in body fluids, the involvement in pathological processes, the demonstrated differential expression in solid tumors and hematological malignancies, and their power to discriminate cancer patients from healthy controls open the prospect for development of minimally invasive, tRF/tiRNA-based biomarker tests. For example, the tRF profiles have been shown to discriminate triple-negative, triple positive breast cancer cells from the normal controls in unsupervised clustering [15] (Fig. 7B,C). The ratio of tRFs/tiRNAs has also been demonstrated as a good indicator of cancer progression-free survival and a candidate prognostic marker [16].
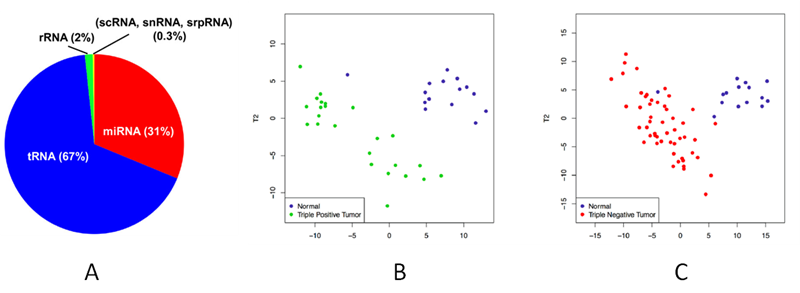
Figure 7. The abundant presence of tRNA derived fragments in serum [17] (A). tRF profiles discriminate triple-negative (B), triple positive breast cancer cells (C) from the normal controls in unsupervised clustering.
How to study tRFs/tiRNAs
nrStar™ tRF&tiRNA PCR Array systematically profiles the abundance and expression of tRFs/tiRNAs. With the wealth data and information, the differentially expressed tRFs & tiRNAs can be identified for the next step studies with the approaches and applications (Fig. 8). Many well established methodologies are similar to what are used for miRNAs, for example, qPCR confirmation, knockdown by LNA oligos, and gain of function studies by synthetic small RNA mimics.
Figure 8. Roadmap for the next step tRF&tiRNA studies.
Related Products
nrStar™ tRNA PCR Array
nrStar™ tRF&tiRNA PCR Array
References
1.Anderson P. and P. Ivanov (2014) “tRNA fragments in human health and disease.” FEBS Lett. 588(23):4297-304 [PMID: 25220675]
2.Zheng L.L. et al. (2016) “tRF2Cancer: A web server to detect tRNA-derived small RNA fragments (tRFs) and their expression in multiple cancers.” Nucleic Acids Res. [PMID: 27179031]
3.Pliatsika V. et al. (2016) “MINTbase: a framework for the interactive exploration of mitochondrial and nuclear tRNA fragments.” Bioinformatics [PMID: 27153631]
4.Cole C. et al. (2009) “Filtering of deep sequencing data reveals the existence of abundant Dicer-dependent small RNAs derived from tRNAs.” RNA 15(12):2147-60 [PMID: 19850906]
5.Burroughs A.M. et al. (2011 Jan-Feb) “Deep-sequencing of human Argonaute-associated small RNAs provides insight into miRNA sorting and reveals Argonaute association with RNA fragments of diverse origin.” RNA Biol 8(1):158-77 [PMID: 21282978]
6.Haussecker D. et al. (2010) “Human tRNA-derived small RNAs in the global regulation of RNA silencing.” RNA 16(4):673-95 [PMID: 20181738]
7.Maute R.L. et al. (2013) “tRNA-derived microRNA modulates proliferation and the DNA damage response and is down-regulated in B cell lymphoma.” Proc. Natl. Acad. Sci. U.S.A. 110(4):1404-9 [PMID: 23297232]
8.Venkatesh T. et al. (2016) “tRFs: miRNAs in disguise.” Gene 579(2):133-8 [PMID: 26743126]
9.Gebetsberger J. et al. (2012) “tRNA-derived fragments target the ribosome and function as regulatory non-coding RNA in Haloferax volcanii.” Archaea 2012:260909 [PMID: 23326205]
10.Sobala A. and G. Hutvagner (2013) “Small RNAs derived from the 5′ end of tRNA can inhibit protein translation in human cells.” RNA Biol 10(4):553-63 [PMID: 23563448]
11.Goodarzi H. et al. (2015) “Endogenous tRNA-Derived Fragments Suppress Breast Cancer Progression via YBX1 Displacement.” Cell 161(4):790-802 [PMID: 25957686]
12.Emara M.M. et al. (2010) “Angiogenin-induced tRNA-derived stress-induced RNAs promote stress-induced stress granule assembly.” J. Biol. Chem. 285(14):10959-68 [PMID: 20129916]
13.Saikia M. et al. (2014) “Angiogenin-cleaved tRNA halves interact with cytochrome c, protecting cells from apoptosis during osmotic stress.” Mol. Cell. Biol. 34(13):2450-63 [PMID: 24752898]
14.Chen Q. et al. (2016) “Sperm tsRNAs contribute to intergenerational inheritance of an acquired metabolic disorder.” Science 351(6271):397-400 [PMID: 26721680]
15.Telonis A.G. et al. (2015) “Dissecting tRNA-derived fragment complexities using personalized transcriptomes reveals novel fragment classes and unexpected dependencies.” Oncotarget 6(28):24797-822 [PMID: 26325506]
16.Olvedy M. et al. (2016) “A comprehensive repertoire of tRNA-derived fragments in prostate cancer.” Oncotarget [PMID: 27015120]
17.Dhahbi J.M. et al. (2013) “5′ tRNA halves are present as abundant complexes in serum, concentrated in blood cells, and modulated by aging and calorie restriction.” BMC Genomics 14:298 [PMID: 23638709]
18.Schageman J. et al. (2013) “The complete exosome workflow solution: from isolation to characterization of RNA cargo.” Biomed Res Int 2013:253957 [PMID: 24205503]
19.Hanada T. et al. (2013) “CLP1 links tRNA metabolism to progressive motor-neuron loss.” Nature 495(7442):474-80 [PMID: 23474986]
20.Martens-Uzunova E.S. et al. (2013) “Beyond microRNA–novel RNAs derived from small non-coding RNA and their implication in cancer.” Cancer Lett. 340(2):201-11 [PMID: 23376637]
21.Lee Y.S. et al. (2009) “A novel class of small RNAs: tRNA-derived RNA fragments (tRFs).” Genes Dev. 23(22):2639-49 [PMID: 19933153]
22.Sharma U. et al. (2016) “Biogenesis and function of tRNA fragments during sperm maturation and fertilization in mammals.” Science 351(6271):391-6 [PMID: 26721685]
23.Greenway M.J. et al. (2006) “ANG mutations segregate with familial and ‘sporadic’ amyotrophic lateral sclerosis.” Nat. Genet. 38(4):411-3 [PMID: 16501576]
24.van Es M.A. et al. (2011) “Angiogenin variants in Parkinson disease and amyotrophic lateral sclerosis.” Ann. Neurol. 70(6):964-73 [PMID: 22190368]
25.Blanco S. et al. (2014) “Aberrant methylation of tRNAs links cellular stress to neuro-developmental disorders.” EMBO J. 33(18):2020-39 [PMID: 25063673]
26.Selitsky S.R. et al. (2015) “Small tRNA-derived RNAs are increased and more abundant than microRNAs in chronic hepatitis B and C.” Sci Rep 5:7675 [PMID: 25567797]