
On average, there are about three m6A modifications per mRNA [1]. Most m6A modified mRNAs contain only a single m6A site, while some mRNAs can have up to 20 or more [2-5]. A “single-m6A site” has no other m6A sites nearby, either up- or down-stream. For multiple m6A sites, adjacent m6A sites tend to aggregate within short regions [6]. A “polymethylated m6A region” is where multiple individual m6A sites are located close together (< 20 nt apart). When a single m6A site and a polymethylated region are contained within 500 nt and the distance between them is < 100 nt, they are merged as one “clustered m6A site”. Single and polymethylated m6A modifications have functional significance for molecular, cellular and organismal phenotypes.
Functional significance of single-m6A sites for cellular and organismal phenotypes
Most m6A-modified mRNAs contain only a single m6A site [5]. The single-m6A modifications are often related to molecular functions in mRNA translation initiation or translation elongation dynamics. They also regulate noncoding RNA decay and activities.
The single-m6A site functions in Cap-independent translation initiation of mRNA
A single-base m6A modification at A103 position in the 5’UTR of Hsp70 mRNA is necessary and sufficient to promote its noncanonical cap-independent translation (Fig. 1) [7]. Under stress conditions, nuclear localized YTHDF2 reader protein protects the m6A from demethylation by FTO. Translation initiation factor (eIF3) can directly bind to the m6A103 and initiate the internal translation, thereby allowing cap-independent translation of HSP70 at a time when the global cap-dependent translation is shutting down in the stressed cells.
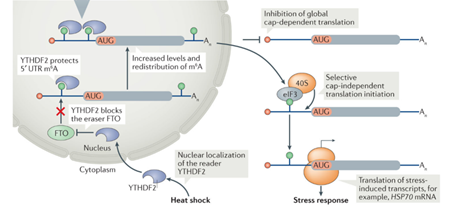
Figure 1. Molecular mechanism of cap-independent translation of HSP70 mRNA under heat shock stress by a single-base m6A modification [7].
The single-m6A site affects mRNA translation dynamics
A single-base m6A modification in mRNA coding sequence (CDS) can affect the translation elongation dynamics [8]. An m6A in a codon can slow down the cognate-tRNA decoding and also act as a barrier/controller for tRNA accommodation and translation elongation. The m6A stalled translation may produce truncated proteins and have chain effects on co-translational protein folding, chaperoning, and localization signal recognition, all of which can potentially drive the protein for different fates or activities [9]. Conceivably, if m6A recodes a codon for another amino acid, it could lead to protein variants with an altered function and substantial biological consequence even at low occupancy [8].
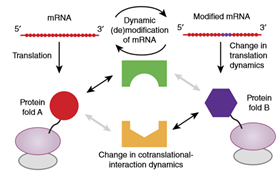
Figure 2. The changed translation elongation dynamics can influence co-translational nascent protein folding or interaction with other partnering factors [8].
The single-m6A site regulates lncRNA decay
Single-m6A sites in lnc-Dpf3 are read by YTH-DF2, which targets the lncRNA for degradation [10]. Chemokines activated CCR7 receptor activated by chemokines removes the m6As in lnc-Dpf3 and prevent its m6A-dependent RNA degradation. The up-regulated lnc-Dpf3 binds to and inhibits HIF1a, thereby suppressing dendritic cell migration and inflammatory response (Fig. 3).
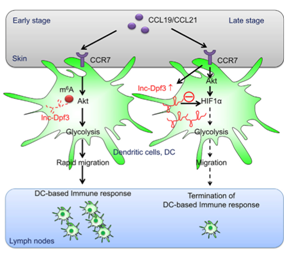
Figure 3. Single-base m6A modifications in lnc-Dpf3 targets the RNA for degradation. Under chemokine stimulation, the m6As are removed and the lnc-Dpf3 is stabilized. The increased lnc-Dpf3 inhibits HIF1a to suppress dendritic cell based immune response [10].
In another lncRNA example, modification at single-base m6A sites (A917, A1025 and A1056) in linc1281 is required to decoy let-7 family miRNAs (Fig. 4) [11]. By sequestering these pluripotency-related miRNAs, the m6A modified linc1281 ensures the cells of mESC identity.
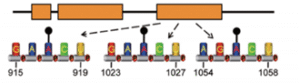
Figure 4. m6A methylation of single-m6A sites in the last exon of linc1281 is required for the lncRNA to bind let-7 miRNAs and act as a competing endogenous RNA (ceRNA) to regulate mESC differentiation [11].
The single-m6A site in 28S rRNA affects cell proliferation and impacts tumor growth
The single-base m6A site in 28S rRNA at A4220 can be modified by a newly discovered m6A methyltransferase ZCCHC4. The m6A4220 modification is required for optimal global translation activity of the ribosome, which affects cell proliferation and impacts tumor growth [12].
Functional significance of polymethylated and clustered m6A regions for cellular and organism phenotypes
Multiple m6A residues packed in clusters are most prominent in mRNA degradation to shape the half-life of mRNAs in cytoplasm. The mRNA metabolic fates are destined by m6A readers targeting them into cytoplasmic compartments by phase separation. Polymethylated regions are also the substrates for heavy m6A modification required for lncRNA activities.
Polymethylated or clustered m6As partition phase separation of mRNAs into subcellular compartments
Polymethylated or clustered m6As, but not singly methylated m6A, can partition the mRNA population into different subcellular compartments via liquid phase separation (Fig.5) [13]. Cytosolic YTH-DF1, 2 and 3 reader proteins spontaneously partition themselves into liquid phases both in vitro and in cells. Polymethylated mRNAs act as a multivalent scaffold for YTHDF binding, which are carried into different phase-separated subcellular compartments, such as P-bodies, stress granules or neuronal RNA granules for different metabolic fates. For example, polymethylated mRNAs stored in stress granules are translationally repressed. Polymethylated mRNAs targeted to P-bodies by YTH-DF2 are degraded [14]. A singly methylated m6A does not have sufficient YTHDF binding affinity for this to occur.
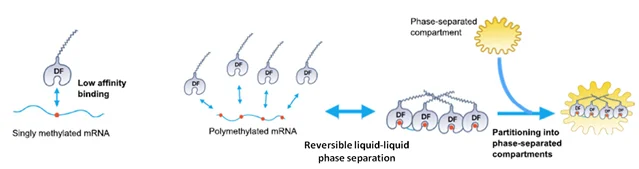
Figure 5. Polymethylated m6As bind cytosolic YTH-DF1,2,3 readers at multivalent affinity and are separated into subcellular compartments for the metabolic fates. A single m6A does not have enough binding affinity for this to occur [13].
Polymethylated m6As regulate mRNA stability
In MYC mRNA, a 250-nt Coding Region Instability Determinant (CRD) immediately 3’ to the CDS harbors multiple m6A sites, which is recognized by IGF2BP1, 2, and 3 reader proteins that promote mRNA stability and translation, thereby globally increasing the gene expression output (Fig. 6) [15].
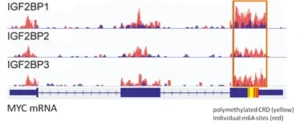
Figure 6. IGF2BP reader proteins bind to polymethylated m6A sites in Coding Region of Instability Determinant (CRD) which stabilize MYC mRNA for increased MYC protein expression.
A region containing polymethylated m6As is conserved for X chromosome silence
lncRNA-XIST is heavily modified at 78 m6A sites — more than any other RNA (Fig. 7). In particular, the “A repeats” region evolutionarily conserved for X-chromosome inactivation activity contains polymethylated m6A sites. m6A modification is necessary for YTH-DC1 reading and recruiting chromatin modifiers for the transcriptional silencing of X chromosome [16].
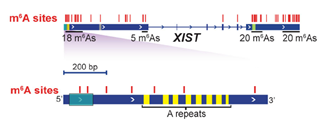
Figure 7. The 78 m6A sites in lncRNA-XIST (red lines, top) and the zoomed-in details of polymethylated m6A sites in the 5’ region (bottom). The evolutionarily conserved “A repeats” (yellow) are essential for the initiation of X-chromosome inactivation by XIST [16].
Related Service
m6A Single Nucleotide Array Service
References
[1] Perry, R. P., et al. (1975) “The methylated constituents of L cell messenger RNA: evidence for an unusual cluster at the 5′ terminus” Cell 4(4):387-94 [PMID: 1168101]
[2] Meyer, K. D., et al. (2012) “Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons” Cell 149(7):1635-46 [PMID: 22608085]
[3] Dominissini, D., et al. (2012) “Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq” Nature 485(7397):201-6 [PMID: 22575960]
[4] Linder, B., et al. (2015) “Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome” Nat Methods 12(8):767-72 [PMID: 26121403]
[5] Zaccara, S., et al. (2019) “Reading, writing and erasing mRNA methylation” Nat Rev Mol Cell Biol [PMID: 31520073]
[6] Zhang, Z., et al. (2019) “Single-base mapping of m(6)A by an antibody-independent method” Sci Adv 5(7):eaax0250 [PMID: 31281898]
[7] Zhou, J., et al. (2015) “Dynamic m(6)A mRNA methylation directs translational control of heat shock response” Nature 526(7574):591-4 [PMID: 26458103]
[8] Choi, J., et al. (2016) “N(6)-methyladenosine in mRNA disrupts tRNA selection and translation-elongation dynamics” Nat Struct Mol Biol 23(2):110-5 [PMID: 26751643]
[9] Kim, S. J., et al. (2015) “Protein folding. Translational tuning optimizes nascent protein folding in cells” Science 348(6233):444-8 [PMID: 25908822]
[10] Liu, J., et al. (2019) “CCR7 Chemokine Receptor-Inducible lnc-Dpf3 Restrains Dendritic Cell Migration by Inhibiting HIF-1alpha-Mediated Glycolysis” Immunity 50(3):600-615 e15 [PMID: 30824325]
[11] Yang, D., et al. (2018) “N6-Methyladenosine modification of lincRNA 1281 is critically required for mESC differentiation potential” Nucleic Acids Res 46(8):3906-3920 [PMID: 29529255]
[12] Ma, H., et al. (2019) “N(6-)Methyladenosine methyltransferase ZCCHC4 mediates ribosomal RNA methylation” Nat Chem Biol 15(1):88-94 [PMID: 30531910]
[13] Ries, R. J., et al. (2019) “m(6)A enhances the phase separation potential of mRNA” Nature 571(7765):424-428 [PMID: 31292544]
[14] Wang, X., et al. (2014) “N6-methyladenosine-dependent regulation of messenger RNA stability” Nature 505(7481):117-20 [PMID: 24284625]
[15] Huang, H., et al. (2018) “Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation” Nat Cell Biol 20(3):285-295 [PMID: 29476152]
[16] Patil, D. P., et al. (2016) “m(6)A RNA methylation promotes XIST-mediated transcriptional repression” Nature 537(7620):369-373 [PMID: 27602518]